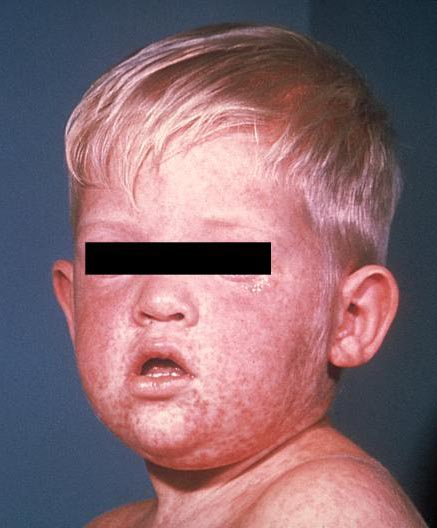
 |
| Uma criança com sarampo. Fonte: https://www.cdc.gov/measles/ |
Se alguém em 2009 dissesse que em dez anos o sarampo voltaria aos noticiários no mundo todo, seria considerado ousado, talvez desinformado ou, na pior das hipóteses, louco. Apesar de surtos ocasionais em um ou outro país, a incidência global da doença estava diminuindo. No ano 2000 os EUA se declararam livres da transmissão do sarampo, e o último caso de sarampo ocorrido de forma autóctone nas Américas havia ocorrido em 2002. Ou seja, os único casos ocorridos a partir de então tiveram origem no exterior e haviam sido trazidos para o continente americano por viajantes. Outros países estavam caminhando para sua eliminação, e já se falava em erradicar a doença nas décadas seguintes. Mas algo deu muito errado no meio do caminho, e cá estamos nós, em 2019, testemunhando uma epidemia de sarampo na maior cidade do Brasil - depois de episódios similares em cidades de Roraima, Amazonas e Pará. A história do ressurgimento do sarampo no mundo é uma parábola do nosso fracasso como civilização. Fracasso em realizar um esforço global para eliminar uma doença que só existe em humanos. Fracasso em manter uma cobertura vacinal adequada em regiões que já não possuem casos da doença. E fracasso em conter a disseminação de outra epidemia, a da mentira e da desinformação nos tempos atuais em que a notícia (incluindo a mentirosa) avança mais rápido do que nosso raciocínio e nosso senso crítico podem dar conta. Pois comecemos do começo: o que é o sarampo, de onde ele veio e o que ele faz?
O sarampo é causado por um vírus da família dos paramixovírus, cujo material genético é feito de RNA. A
cepa ou variedade de sarampo que circula nos dias de hoje é um parente próximo do vírus da peste bovina
(Rinderpest, uma doença que foi erradicada no início do século XXI graças
à vacinação do gado), e relógios moleculares indicam que a separação entre as
duas linhagens provavelmente ocorreu há cerca de mil anos.
Nesse período, a circulação do vírus que daria origem ao sarampo entre humanos favoreceu mutações que o
tornaram mais especializado, a ponto de ele perder sua afinidade com o gado e
circular apenas entre seres humanos. Considerando uma margem de erro de alguns
séculos, essa datação condiz com a primeira descrição do sarampo como doença
distinta da varíola e da catapora, feita pelo médico persa Rhazes em Bagdá por
volta de 910 d.C. É possível que outras cepas do
ancestral do sarampo e da peste bovina tenham sido transmitidas para humanos ou
até mesmo circulado de um humano para outro diversas vezes nos séculos ou
milênios anteriores, mas nesses casos o surto não foi à frente porque acometeu
populações isoladas e minguou antes que um desses indivíduos doentes
transmitisse o vírus para outras pessoas. A variedade atual de sarampo se
espalhou com facilidade graças à maior densidade populacional existente no
Oriente Médio e depois na Europa, no norte da África e no restante do planeta
naquela época, bem como à rede comercial e de estradas que permitia o
deslocamento relativamente rápido entre vilarejos. Colaborou para isso também o
fato de o sarampo ser uma doença altamente contagiosa e poder ser transmitida
pelo ar até mesmo três a cinco dias antes de o indivíduo doente iniciar os
sintomas. Em um ambiente fechado, o vírus pode permanecer suspenso no
ar por até duas horas. Dessa forma, um indivíduo com sarampo consegue
transmitir a doença para até dezoito pessoas,
e quando ele começar a manifestar os sintomas outras pessoas já entraram em
contato com o vírus. É uma das doenças mais contagiosas que a Humanidade conhece.
E que sintomas são esses? O sarampo é ao mesmo tempo uma doença respiratória (como a gripe e o resfriado) e exantemática (ou seja, causadora de lesões avermelhadas na pele, como a rubéola e a catapora). Os primeiros sintomas são febre (geralmente alta, acima de 39°C), tosse, coriza, irritação nasal e conjuntivite - ou seja, vermelhidão nos olhos. Nessa fase da doença podem surgir lesões esbranquiçadas na face interna das bochechas, visíveis ao exame da cavidade oral, chamadas manchas de Koplik e que são consideradas patognomônicas do sarampo - ou seja, nenhuma outra doença conhecida é capaz de causar o mesmo sinal.
 |
| Manchas de Koplik na mucosa oral. Fonte: https://medlineplus.gov/ency/imagepages/2558.htm |
Em alguns dias surgem lesões avermelhadas na pele, inicialmente na cabeça e se espalhando depois para o tronco e os membros. O aspecto é similar ao de outras doenças exantemáticas, ou ao de uma reação alérgica. Outros sintomas mais atípicos podem surgir em alguns pacientes, como infecções de ouvido ou diarreia. Em geral, eles ocorrem mais em crianças pequenas, com menos de 2 anos de idade.
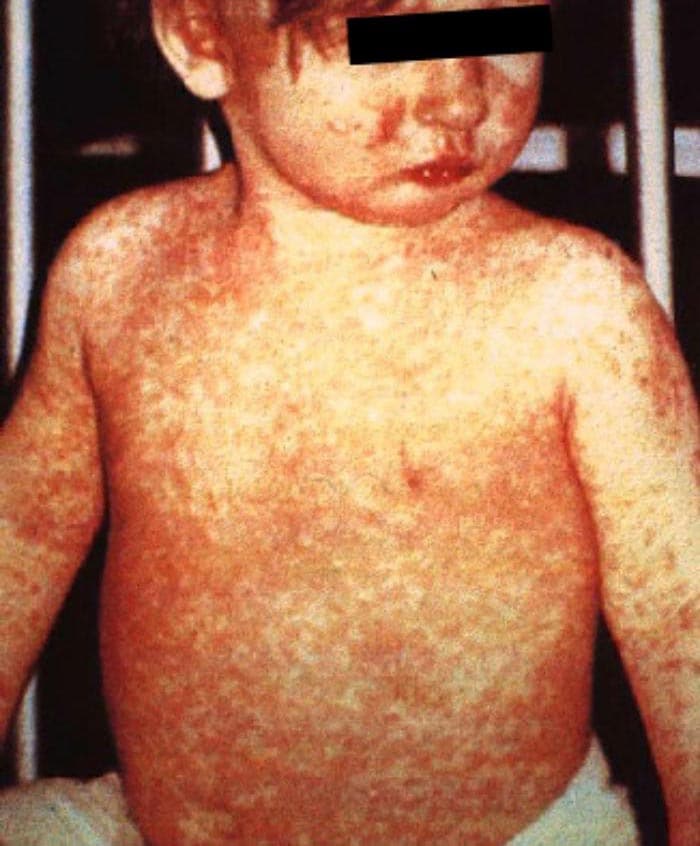 |
| Exantema causado pelo sarampo. Fonte: https://www.cdc.gov/measles/images/people_measles7.jpg |
Na grande maioria dos casos a pessoa - geralmente uma criança - se recupera sem sequelas. No entanto, 1 a cada 5 pessoas com sarampo precisa de internação (nos EUA, embora seja possível que no Brasil as estatísticas também girem em torno disso). 1 em cada 1000 crianças pode desenvolver encefalite (acometimento do cérebro pelo sarampo), que pode ser fatal ou resultar em sequelas para o resto da vida. Pneumonias, sejam causadas pelo próprio sarampo ou por bactérias que aproveitam a mucosa respiratória já lesada para causar infecção, ocorrem em 1 a cada 20 pacientes com sarampo, e podem também levar à morte. 1 a 3 em cada 1000 crianças acaba morrendo de complicações respiratórias ou neurológicas - um número que chegava a 30% quando ainda não havia antibióticos. E existe uma entidade clínica mais rara chamada pancencefalite esclerosante subaguda, que pode acometer crianças anos após o contato com o vírus e levar a distúrbios cognitivos e de comportamento, degeneração neurológica e óbito.
Em geral, o sarampo é mais grave em crianças menores de 5 anos, adultos com mais de 20 anos, gestantes, pessoas desnutridas (principalmente aquelas com baixos níveis de vitamina A) e pessoas com imunidade baixa - incluindo pacientes com AIDS com quantidades de linfócitos CD4 menores que 200, segundo o CDC. Em crianças desnutridas, a mortalidade do sarampo pode chegar a assustadores 10%.
Por essa facilidade de se disseminar e causar surtos e
epidemias e pela possibilidade de levar a uma pneumonia ou a uma inflamação no
sistema nervoso central, o sarampo foi durante muitos séculos uma doença
mortal, especialmente em crianças. É bom lembrar que, embora pareçam pequenos, esses números (1 caso de encefalite a cada 1000 pacientes, 1 a 3 óbitos a cada 1000 pacientes...) ficam muito altos em termos absolutos quando estamos falando de populações maiores. Uma cidade com 1 milhão de habitantes terá 1 a 3 mil óbitos por sarampo se a mortalidade permanecer dentro dessa estatística.
Infelizmente não existe uma medicação capaz de curar o sarampo. Diversos antivirais já foram testados, mas nenhum se mostrou capaz de conter o vírus ou suas consequências. Devido à correlação entre deficiência de vitamina A e casos mais graves, tem sido comum ofertar vitamina A para os pacientes com sarampo. De resto, o que se pode fazer é oferecer suporte: conter a febre, tratar pneumonias bacterianas e aguardar a recuperação.
A única medida efetiva contra o sarampo é a vacina. A primeira versão surgiu na década de 1960, e desde então ela tem sido adotada no mundo todo. Seu sucesso em conter epidemias e surtos é inegável. A versão atual utilizada no Brasil é composta por vírus atenuados (ou seja, viáveis porém fracos o suficiente para não causar doença a não ser que a pessoa esteja com a imunidade muito debilitada). Em geral, vacinas de vírus atenuados tendem a garantir uma imunidade maior e mais prolongada do que aquelas causadas por vírus mortos. É administrada juntamente com as vacinas de caxumba e rubéola na mesma ampola - daí seu nome MMR - sigla em inglês para Mumps (caxumba), Measles (sarampo) and Rubella (rubéola). No Brasil, a mortalidade despencou de 5% para zero no último quartil do século XX. A última epidemia de sarampo havia ocorrido em 1997, e os casos da doença caíram dramaticamente desde então. Declaramos que o país estava livre da doença, e que novos casos só ocorreriam se viessem de fora do país. Em todo o mundo, as mortes anuais por sarampo caíram de 1,2 milhão em 1985 para menos de 100 mil em 2016 graças às campanhas de vacinação. Falava-se em erradicar a doença nas décadas seguintes, assim que conseguíssemos fazer os esforços mundiais penetrarem nos rincões da Ásia e da África. O sarampo (e talvez a poliomielite) se juntariam à varíola e à peste bovina, as duas outras doenças que conseguimos eliminar da face da Terra. O clima era de otimismo.
No entanto, começamos a relaxar. A cobertura vacinal caiu. Parte da culpa foi dos governos e das autoridades de saúde locais, que não aplicaram as duas doses recomendadas (idealmente aos 12 e aos 15 meses de vida) como deveriam aplicar. Por que se preocupar, já que é uma doença que está desaparecendo? Mas parte da culpa também era das pessoas. Para que vacinar meus filhos contra uma doença que está desaparecendo? Por que vacinar se o meu vizinho já se vacinou e não vai transmitir para mim nem para meus filhos? As gerações mais novas já não sabiam o que era sarampo. Já não se importavam. E com isso, a estratégia de sucesso do controle do sarampo pela vacina, o efeito rebanho, se tornou insustentável.
Explico-me: como o sarampo é uma doença extremamente contagiosa, uma pessoa com a doença disseminará o vírus para todos aqueles em volta desde três a cinco dias antes do início dos sintomas até quatro dias depois do surgimento das lesões de pele, em média. Para evitar que essas pessoas em volta contraiam a doença, é preciso que elas estejam vacinadas. No entanto, existem alguns grupos de pessoas que não podem receber a vacina, porque seu sistema imunológico está debilitado demais para lidar com o vírus atenuado que ela contém. Gestantes (não por si mesmas, mas pelo feto que não possui sistema imunológico maduro), bebês até 6 meses de idade e pessoas com doenças que afetam a imunidade (AIDS com CD4 menor que 200, receptores de transplantes, pessoas com neoplasias como leucemia ou que têm alguma doença genética que resulta em baixa imunidade) não podem ser vacinadas, e por isso estão em risco de adoecer se entrarem em contato com alguém com a doença. Agora imagine um bairro com cem casas, cada uma habitada por uma pessoa. Em uma ponta do bairro vive uma pessoa com sarampo. Em outra mora uma pessoa que não pode tomar a vacina - digamos, uma pessoa que trata uma leucemia. É pouco provável que a pessoa com sarampo interaja com o indivíduo com leucemia a ponto de transmitir a doença para ele, portanto a transmissão será muito mais fácil de ocorrer se "saltar" de uma pessoa para outra. Ou seja, se o indivíduo com sarampo transmitir a doença para um de seus vizinhos mais próximos, e esse adoecer e transmitir para outro, que transmitirá para outro, e assim por diante até que o vizinho do lado do paciente com leucemia também esteja com a doença. É o que aconteceria se nenhum dos moradores se vacinasse. No entanto, se todas as outras 98 pessoas do bairro tiverem sido vacinadas (excetuando o paciente com sarampo e o com leucemia), nenhuma delas contrairá a doença e portanto não poderá transmitir o vírus adiante. Porém, se 50% delas não tiver sido vacinada, a transmissão ainda será muito provável. Modelos matemáticos indicam que é preciso que pelo menos 95% das pessoas estejam imunizadas para que o risco de transmissão seja desprezível, e essa era a meta das campanhas de vacinação: vacinar 95% das pessoas. 95% das pessoas imunizadas no bairro, no metrô, no elevador do prédio de escritórios, no shopping... Se essa proporção fosse garantida, jamais teríamos novas epidemias de sarampo. Mesmo que por azar uma pessoa transmitisse a doença para outra que não estivesse vacinada, a chance de que uma das duas vítimas transmitisse para uma terceira suscetível ao redor era quase nula.
No entanto, vacinar 95% das pessoas não é uma tarefa fácil. Exige um esforço intenso e contínuo para correr atrás inclusive daquelas que não procuraram as unidades de saúde. É caro e demanda esforço, recursos e vontade do poder público. Nem sempre temos os três juntos. E com a diminuição no número de casos, a cobertura vacinal começou a minguar. Para ser justo, no Brasil dificilmente chegou aos 95%, mas se manteve em torno dos 90% em grande parte do território. Mas depois caiu para menos de 90%. Depois para menos de 80%. Depois para menos de 70% em algumas regiões. No entanto, não se pode culpar apenas a negligência do poder público, porque casos novos de sarampo começaram a ressurgir em outros lugares do mundo. Nesse caso, foi por culpa de outra epidemia: a da ignorância.
Em 1998, um médico britânico chamado Andrew Wakefield publicou um estudo com 12 pacientes em que dizia ter encontrado uma relação entre a vacina MMR e o autismo. Segundo ele, o vírus vacinal permanecia nos tecidos linfoides do intestino, deflagrava uma resposta imunológica anormal e isso resultava em autismo. Apesar da pequena amostra (12 pacientes é um número muito pequeno para se tirar conclusões, na grande maioria das vezes) e de outras inconsistências, o artigo foi aceito e publicado no Lancet, uma das revistas médicas mais conceituadas do mundo. O que Wakefield não revelou foi que ele recebera financiamento de advogados que tinham como objetivo processar os produtores de vacinas em nome de seus clientes - pais cujos filhos haviam sido diagnosticados como autistas pouco tempo depois de receber a vacina. Também não revelou que ele próprio estava patenteando uma vacina própria que, segundo ele, não causaria autismo. Ou seja, havia conflito de interesses na publicação de Wakefield, que aparentemente esperava lucrar com a informação contida em seu estudo, tanto com o patrocínio de advogados quanto com a patente da vacina nova. Novos estudos foram realizados, com um número muito maior de participantes, e nenhum deles encontrou absolutamente nenhuma relação entre a vacina MMR e o autismo. Descobriu-se depois que alguns daqueles 12 pacientes do estudo já exibiam sinais sugestivos de autismo antes de tomar a vacina, e outros haviam sido diagnosticados de forma errada. No fim das contas, a publicação de Andrew Wakefield ficou caracterizada como fraude, e o Lancet a retirou de sua revista após 12 anos (o mais intrigante é como e por que motivo ele conseguiu ser publicado, evidenciando que não é porque um artigo está em uma revista bem-conceituada que ele necessariamente é confiável). Wakefield perdeu o direito de exercer a Medicina no Reino Unido e nos EUA, onde vive hoje.
No entanto, o estrago estava feito. A notícia falsa aparece na manchete, e a retratação dizendo que ela é mesmo falsa aparece na página seis. Explicações fáceis e simples chamam muito mais atenção e atraem muito mais pessoas, especialmente quando estamos falando de uma doença cuja causa é até hoje desconhecida. Sabe-se que o autismo tem algum componente genético e algum ambiental, mas ainda não está clara a influência de cada um nem quais são esses componentes especificamente - o que se pode afirmar, com segurança, é que a vacina não está entre eles. O transtorno de espectro autista (porque vai desde quadros leves como a síndrome de Asperger até casos em que a pessoa é incapaz de falar) parece ter início nos primeiros meses após o nascimento ou mesmo intraútero, mas seus sinais começam a ficar mais claros após o primeiro ano de vida quando a criança começa a falar e a interagir mais socialmente - e justamente na mesma época em que as crianças recebem a vacina MMR. Mas nada disso conseguia convencer milhares ou milhões de pais de crianças autistas, ansiosos por um culpado para a dificuldade de interação social dos seus filhos. Culpar a vacina e dizer que Wakefield era vítima de uma conspiração global era muito mais fácil do que se conformar com o fato de que ainda não sabemos o que causa o transtorno de espectro autista. Desde a publicação fraudulenta de Wakefield, a procura pela MMR mesmo em países desenvolvidos despencou. Bairros de Los Angeles, Nova York e Londres passaram a ter proporcionalmente menos crianças vacinadas contra o sarampo do que comunidades isoladas no interior da África. Culpou-se o timerosal, o conservante à base de mercúrio adicionado a algumas vacinas (mas não à MMR) que fazia com que durassem mais tempo - e que também estava presente no Merthiolate até sua fórmula mudar. Mesmo que uma dose de vacina com timerosal tivesse menos mercúrio do que uma lata de atum em conserva, o timerosal foi banido - e levar vacinas às regiões remotas ficou mais caro. Depois, quando ficou provado que o timerosal não era o culpado, culparam o alumínio, presente em pequenas doses das vacinas, que novamente se descobriu que não tinha nenhuma relação com autismo. Os negadores da ciência continuam em busca de um culpado pelo autismo, e vão se agarrar ao primeiro que for apontado por um charlatão qualquer. Wakefield pode não ser autorizado a praticar a Medicina nos EUA, mas está ganhando rios de dinheiro com o movimento antivacinas, graças a seus seguidores descerebrados.
Com a queda na procura pela vacina, novos casos de sarampo voltaram a surgir no mundo todo. Alguns foram motivados pela desconfiança semeada por Wakefield sobre a vacina, como em muitos lugares da Europa e dos EUA. Outros, pelo colapso de sistemas de saúde locais devido a guerras ou crises econômicas, como na Ucrânia e na Venezuela. E outros também pela recusa em vacinar, mas vindo de grupos religiosos ultraortodoxos que proíbem vacinas entre seus pares.
O Brasil, livre do sarampo por anos, estava despreparado para o retorno do vírus. Com o colapso econômico da Venezuela sob Nicolás Maduro, a cobertura vacinal no país despencou, e teve início uma epidemia no país a partir de 2017. Com a crise econômica, milhares de venezuelanos fugiram do país. Um pequeno número veio para o Brasil (alguns milhares, mas nada comparável ao número de refugiados na Colômbia ou no Peru, por exemplo), e com o vírus circulando inicialmente entre os venezuelanos, não tardou para que fosse transmitido também para os brasileiros. Surtos irromperam em Boa Vista, Manaus e Belém a partir do início de 2018. Importante destacar: não se pode fugir da responsabilidade e culpar os venezuelanos pela epidemia de sarampo entre os brasileiros. Se tivéssemos nossa cobertura vacinal nos 95% recomendados, a epidemia vista no Norte do Brasil jamais teria acontecido. Com vacinação de bloqueio em todos aqueles que tiveram contato com casos, a epidemia foi contida - não sem antes causar mais de dez mil vítimas, com doze óbitos, a maior parte no Amazonas e em Roraima.
No entanto, vivemos agora outra epidemia, por casos originados não na Venezuela, mas sim em países da Europa e em Israel. No caso do indivíduo que trouxe o vírus de Israel, provavelmente há ligação com um surto da doença no país iniciada em comunidades de judeus ultraortodoxos que se recusam a se vacinar e a vacinar seus filhos. Não por acaso, um surto em Nova York também envolveu uma comunidade de judeus ortodoxos. São Paulo apresenta o maior número de casos: são quase 500, 75% deles concentrados na capital. Minas Gerais e Rio de janeiro também registraram casos, bem como o Pará, cujos casos ainda parecem derivar da epidemia que acometeu o norte do Brasil vindo da Venezuela. Os dados mais recentes disponíveis são do dia 12 de julho, e provavelmente mudarão com o avançar da epidemia. Novamente, vale aqui o que valia para o Norte do Brasil em 2018: a epidemia dificilmente estaria ocorrendo se a cobertura vacinal estivesse nos 95% recomendados.
A principal recomendação para controle da epidemia é e continuará sendo a vacinação. Todas as pessoas com mais de 1 ano e menos de 30 anos devem tomar 2 doses da vacina. Geralmente a primeira dose é dada aos 12 meses e a segunda aos 15, mas caso o indivíduo não tenha sido vacinado nessa época ele pode receber a vacina em qualquer idade até os 30 anos, em duas doses separadas por pelo menos 30 dias. Pessoas entre 30 e 49 anos teoricamente entraram em contato com o vírus ou receberam uma dose da vacina em algum momento da vida, então teoricamente só precisam de uma nova dose. Pessoas com 50 anos ou mais em tese já entraram em contato com o vírus do sarampo uma vez na vida e, graças à memória imunológica, não precisam se vacinar. No estado de São Paulo, devido ao crescente número de casos, esse limite entre precisar de uma dose da vacina e não precisar se vacinar foi estabelecido como 60 anos, e não 50. Pessoas que entraram em contato com alguém com suspeita de sarampo devem tomar a vacina preferencialmente em até 72 horas, independentemente de estado vacinal prévio. Ou seja, mesmo se já receberam duas doses - essa estratégia é conhecida como vacinação de bloqueio. Em indivíduos sem contraindicações, tomar uma terceira dose não causará nenhum mal - na dúvida entre vacinar e não vacinar, vacine. As contraindicações continuam as mesmas: gestantes, menores de 6 meses, imunossuprimidos e também pessoas que possuem alergia à vacina. Pessoas que comprovadamente já tiveram sarampo estão imunizadas e não precisam ser vacinadas. Bebês entre 6 e 12 meses de idade podem tomar a vacina para se proteger temporariamente, mas não há garantia de que a imunidade de memória esteja madura a ponto de proteger para o resto da vida. Assim, essa dose feita antes dos 12 meses não contará na carteirinha de vacinação, e eles deverão tomar as duas doses regulares aos 12 e aos 15 meses. Nos casos em que o indivíduo entrou em contato com alguém com sarampo e possui alguma contraindicação à vacina, a recomendação é a aplicação de imunoglobulinas - anticorpos externos que neutralizam o vírus antes que eles tenham efeito sobre o organismo. A distribuição de imunoglobulinas para a população geral é fortemente desencorajada porque elas são muito caras, difíceis de obter e, se aplicadas em massa, podem faltar para aqueles contactantes que realmente precisarem delas. Como medida de proteção coletiva, a vacina é o instrumento recomendado.
As recomendações mudam constantemente, assim como os números da epidemia. Por isso é recomendado acompanhar os informes e as publicações mais recentes. Pode ser que nos próximos dias tudo o que está escrito no parágrafo anterior mude. Também é possível que consigamos conter essa epidemia, ou que ela se espalhe por outros estados. Uma pessoa em um avião fechado em um voo de 3 horas pode transmitir o vírus para praticamente todos os suscetíveis que estavam no mesmo voo, levando o vírus para outros estados e outras cidades. Um indivíduo vindo de São Paulo passou por duas cidades no Espírito Santo a caminho de outra cidade em Minas Gerais, e obrigou a vigilância epidemiológica local a realizar vacinações de bloqueio em todos os contactantes. Outras localidades certamente têm e terão situações similares. Pode ser que consigamos conter essa epidemia, mas que no mês, semestre ou ano seguinte outra pessoa, vindo de outro lugar do planeta, traga outro vírus e dê início a outro surto ou outra epidemia. Em tempos de viagens internacionais que duram horas, a única forma de estar preparado é garantir que a população do destino esteja imunizada. Repetindo o que já escrevi no começo do texto: o sarampo é uma doença extremamente contagiosa e que pode matar.
O sarampo tem como único hospedeiro o ser humano, sobrevive algumas horas no ambiente e quem já teve a doença ou recebeu as doses adequadas da vacina já está imunizado. Eliminá-lo não é impossível, mas depende de esforços conjuntos e da consciência de que, se eu não me vacino, posso prejudicar o vizinho que não pode se vacinar. Acima de tudo, a luta que estamos perdendo contra o sarampo evidencia que a nossa ignorância e nossa inclinação a acreditar em charlatães são a nossa ruína. Só o combate à desinformação pode impedir que cometamos atos de sabotagem contra nós mesmos e nossos filhos, expondo-os a males que já poderiam ser eliminados da face da Terra. A julgar pelo número crescente não só de anti-vaxxers, mas também de terraplanistas e negacionistas climáticos, estamos perdendo feio essa briga.

